A PHYSICIST'S VIEW OF THE USE OF FEEBLE ELECTRIC DIRECT CURRENTS
TO REPAIR TISSUE AND REPLACE BODY PARTS
(PART ONE)
BY GARY WADE, PHYSICIST
In this article I
am going to give a review of the essential aspects and results of Dr. Robert Becker's research group and the
work of others, as was laid out in his book , THE BODY
ELECTRIC , which will supply the solid foundation needed to support a
simple yet critical observation, which explains how in general to regenerate
mammal body tissues, including the spinal cord. (See links to collected articles of Gary Wade at end of this paper.)
Mammals
are lacking a single simple tissue electric direct current "code",
which would allow them to repair server
tissue damage such as an amputation.
After server tissue damage in mammals all of the other needed biological
repair processes are ready and just waiting for this electric direct current
code. Complete tissue repair or
replacement would then come as a natural course of events, following this direct
current code. This direct current
code can be simply and easily artificially supplied after server damage. This direct current generates a physical
chemistry process which has the net result of significantly increasing above
normal the positive ion concentrations
and decreasing the negative chlorine ion concentration below normal in the
saline plasma solution outside the cell membranes in the damaged tissue. These much higher positive ion concentrations
and lower than normal chlorine ion concentrations in combination with nerve
cell and other cell hormone secretions foster tissue repair and or
replacement. Though the direct current
regeneration code can be simply and easily implemented in practice, the current
allopathic medical system is not likely to implement it. The people who need this regeneration process
will probably have to light a "fire" under the hind quarters of
allopathic medicine's ignorant, arrogant, complacent, and corrupt
leadership.
Some
years ago I read the book THE BODY
ELECTRIC by Robert O. Becker,
M.D., and Gary Selden.(1)
For me it was a fascinating book for it dealt with many of the
electrical aspects of biology, which I have a keen interest in. Shortly after reading Becker's book my sister
informed me that a dear friend of her's had become a quadriplegic as a result
of a gun shot to his neck. The
perpetrator of this act had been apprehend shortly after the shooting. It was learned that it had been a case of
mistaken identity. This hired killer had
mistaken Richard for his intended target.
Because of Richard's tragic
situation , I decided to see, if there was something I could do to cure spinal
cord damage and breakage. To do this I
would have to figure out the secret to repairing / replacing body tissue in
mammals. Becker and his associates had
essentially done all the needed experimental work , as was clearly laid out in
his book THE BODY ELECTRIC.
What was required was someone to put all the pieces together into a
coherent approach to regeneration, which explained what was the critical factor
to get tissue regeneration or replacement.
Becker and his fellow researchers had even effectively done this. They had only failed to recognize a positive ion increasing concentration process
and a decreasing negative chlorine ion concentration process caused by the
direct current which triggers cell dedifferentiation in certain cell types necessary for beginning the tissue
regeneration process.
Figure 1A crudely illustrates the Anatomy of the nervous
system of the salamander. Even though
there is a great deal of difference in the fine details between a salamander's
nervous system and that of a mammal, it can still be thought of as a forerunner
to ours (see
Figure 1B). And in fact there is one critical similarity
between salamander and mammal nervous systems which is critical for
regeneration of damaged tissue. That
similarity is that both nervous systems have Schwann type cells which surround
motor nerve axons and sensor nerve fibers, which also transport direct electric
currents to and from body tissue (see Figure 1C and Figure 1D). The
Schwann cells coating the motor nerve fiber axon carry negative charges
(electrons) away from the central nervous system and deposit them into body
tissue in the form of hydroxyl ions (
As we
proceed in this article, it will become clear how thesecurrents are critical to
regeneration. The 2 key physiological difference for us to note in the
nervous system anatomy between a salamander and a mammal is that a salamander
has approximately three to four times more nerve tissue mass per unit volume of
tissue when considering tissue outside the central nervous system. This means that salamanders have potentially
approximately three to four times more current transporting ability to and from
their tissue.(2) However, it
is the basic similarities between the salamander and mammal nervous systems
which initially gave hope, that was later experimentally verified, that a
mammal could regrow body parts, just as a salamander could. It is the astounding regeneration abilities
of salamanders that has kept some researchers so hopeful for mammals ( humans )
to be able to some day regenerate body tissue and parts throughout the entire
body, if only the salamander's secret could be learned. Salamanders can regrow their arms, legs,
tails, half a heart ( after surgical removal ), repair a cut or severed spinal
cord, regrow a removed eye, and regrow large amounts of their brain ( after
surgical removal ).
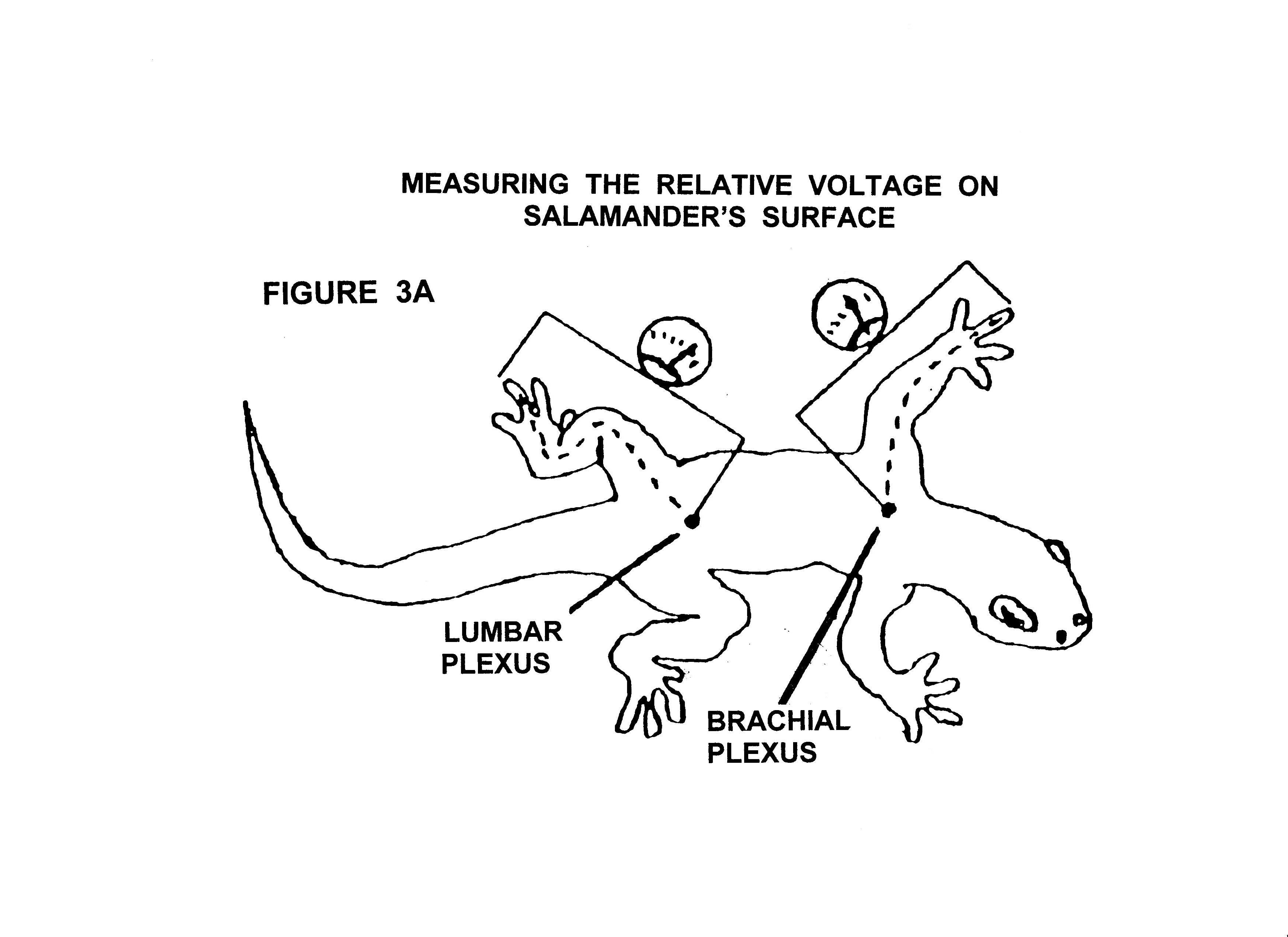
Figure 2 illustrates the qualitative distribution and
polarity of the voltage found on the skin surface of the salamander. The negative areas have a very slight surplus
of electrons and the positive areas a very slight deficiency of electrons. Figure 3A and Figure 3B illustrate
how the relative voltage changes with position on the legs and arms on a
salamander's body relative to the voltage potential at the reference points,
the lumbar plexus and brachial plexus respectfully. In Figure 3B the reference voltage has
arbitrarily been set to zero for graphing purposes.
Figure 4 illustrates a sponge soaked in salt water
mockup of a salamander, which Becker made.
His idea was to consider a salamander's body essentially a saline
solution with current sources imbedded in it.
Becker used the contact potential difference between two dissimilar
metals ( copper and solder ), along with their electrochemical potential
differences to supply the driving voltage which supplied a weak electric
current in the salt water solution of the salamander mockup. The copper charges up negative when in
contact with solder ( lead and tin alloy )
and the solder charges up positive.
The electrically conductive salt solution completes the circuit between
the exposed copper and solder electrodes shown in Figure 4.
Positive ions of lead ( Pb++ ) and tin ( Sn++
) "dissolve" into the salt
solution along with chlorine gas generation and minor positive hydrogen ion
generation and molecular oxygen generation at the positive solder
electrode. Negative hydroxyl ions (
Figure 5A illustrates the re growth / regeneration
process a salamander goes through after it's arm is amputated just above the
elbow. Figure 5B illustrates how the
voltage potential at the wound / amputation site relative to the reference
point ( brachial plexus ) potential , arbitrarily set to zero, changes with
time as the wound site changes into a blastema and grows into a new arm. The final voltage reading on Figure 5B is from the new fingers. It is the salamander's Schwann cells which
supply the observed voltage / current changes associated with
regeneration. The relative voltage
increase at the amputation site indicate a similar increase in negative current
being delivered to the wound area. Becker performed experiments(3)
which showed that the current carrier in the Schwann cell coating is a N - type semiconductor. The N - type semiconductor is presumably the triple - stranded protein
collagen , which is used by the body for cohesion between cells and which is
also a known N - type semiconductor.
A key
experiment was performed by Becker's research team on frog red blood cells
which gives the key information to answer the question: How can cells dedifferentiate from a specific
functional cell type such as a blood cell type into primitive embryonic looking
cells of the blastema. These blastema
cells then divide / multiply and then differentiate under the direction /
control of nerve cell hormone secretions and adjacent cell hormones secreted
into all the cell types needed to form the reforming arm tissue after
amputation, as is illustrated in Figure 5 A. Figure 6A and Figure 6B illustrate
Becker's key experiment carried out on frog red blood cells.(4) Frog and salamander red blood cells unlike
mammal red blood cells, still have their cell nucleus containing a complete
copy of the frog or salamander genetic material ( the chromosomes ). The salamander red blood cells are the main
target cells that the salamander uses to form the embryonic primitive cells of
the blastema. In mammals the target
cells used for forming the primitive looking dedifferentiated cells of the
blastema are apparently mainly the fibroblast cells, Schwann cells, bone marrow cells and glia and ependymal
cells in spinal cord repair. Observing
frog red blood cells with a microscope while exposing them to different values
of minute direct current flow in their surrounding saline medium (Ringer's
solution), the frog red blood cells change into primitive embryonic looking
cells. In Becker's particular
experimental set up ( illustrated in Figure 6A ) he found
current flows between 200 and 700 picoamps ( 10-12 amps ) caused red blood cells to dedifferentiate
into primitive embryonic looking cells.
First the blood cells at the negative electrode dedifferentiated, then
at the positive electrode, and then spreading across the rest of the
chamber. Within a few hours all the
blood cells had become completely unspecialized, lost their hemoglobin and
their nuclei had reactivated.
Since the
direct current in the saline solution can not flow through the red blood cell
interiors do to their bi - lipid cell membranes acting as insulators, Becker's
group proposed an interaction between the
direct current and the cell membrane which released derepressor proteins
on the inside of the cell membrane which derepressed genes and allowed or made
the cell dedifferentiate into a primitive looking cell type. This hypothesis however does not
explain: 1) Why only a narrow minuscule direct current
flow range ( a current window ) works to dedifferentiate cells, 2) Why the dedifferentiation starts first at the
negative electrode region and then the positive electrode region and then to be
followed through out the entire chamber.
I am
going to propose a similar but different hypothesis as to why minuscule current
flow can cause cells to dedifferentiate.
Becker's successful experiment to partially regrow a amputated rat arm
using a implanted negative current source, will be used to illustrate my
hypothesis.
Figure 7A illustrates Becker's partially successful
experimental attempt to regrow a rat's amputated forearm. From experimental data gained from observing
salamanders regrow amputated limbs, Becker was able to estimate the amount of
"negative" direct current flow needed at the amputation site on a
rat's arm to form a blastema and to obtain regeneration or regrowth of the
arm. Becker used the contact potential
difference between two dissimilar metals to supply the needed direct current
flow. This direct current flow would now
be flowing in the saline body fluids of the rat. The two dissimilar metals were platinum and
silver with a carbon high ohmage resistor connected between them to control the
amount of current flow produced. The
device formed this way and illustrated in Figure 7B was
implanted in the rat arm with the negative electrode placed just behind the
amputation site where the blastema is to form.
Figures 7C illustrate the type of physical chemistry
reactions that occur at and between the saline solution and electrode surface
regions with this device. The negative
platinum electrode becomes a continuous hydroxyl ion ( OH- ) source from a set of physical chemical
reactions represented by Equations 1, 2, and 3.
Na+ +
e _______> Na
; Equation 1
Na + H2O ________> NaOH
+ H ;
Equation 2
NaOH __________> Na+ +
Equation
1 indicates the net result of an
electron (e) quantum tunneling ( jumping ) from the platinum metal surface to
the positive potential well of the Na+ ion in its own water molecule complex only a few atomic diameters away from
the platinum metal surface. Equation 2
indicates how the neutral sodium atom formed by the reaction of Equation 1 will quickly react with any nearby water
molecule to form sodium hydroxide ( NaOH ) and a free hydrogen atom. Equation 3 indicates how the sodium hydroxide
is very unstable in water and rapidly decomposes into a sodium ion ( Na+ )
and a hydroxyl ion (
The
sodium ion ( Na+ ) of Equation 1 could be replaced by a potassium
ion ( K+ ) and achieve the same result. The hydroxyl ion (
The
positive electrode has an analogous set of ion reactions occurring which offset
or balance out the charge displacements occurring at the negative
electrode. At the positive electrode
silver ions (Ag+ ) are going into solution repelling the other
positive ions of the saline solution, while at the same time attracting in
negatively charged chlorine ions ( Cl- ) to balance off or counter
the electric field generated by the silver
positively charged ions . Chlorine ions recombine at the positive
electrode forming chlorine gas ( Cl2 ) and donating their extra
electron to the positively charged silver electrode. Also, minor amounts of molecular oxygen and
positive hydrogen ions are being generated at the silver electrode surface by
the reaction indicated in Equation 4.
2
H2O O2 +
4 H+ + 4 e Equation 4
The positive hydrogen ions also draw in negative
chlorine ions to shield the electric field generated by the hydrogen ion
positive charge.
As is
well known from biological research, the type and amount of ion transport
across the cell bi - lipid membrane and
therefore the concentrations of various ion types inside a cell, controls much
of a cell's physiology and genetic activity.
All cell membranes have various types of ion channels for each ion type,
i.e. Na+, K+ , Cl-
, Ca++ , etc.. The various ion concentrations outside the
cell lipid membrane strongly influence the ion transport across the cell membrane
and therefore the ion concentrations inside the cell and thereby cell
physiology and genetic activity.(6,7,8,9,10,11,12,13)
From our
above discussion of how hydroxyl ion generation causes higher than normal
positive ion concentrations around or in the region of a negative electrode we
will now also see how higher than normal positive ion concentrations around a
positive electrode can occur. Consider
the hydroxyl ion generation process , which will occur in the set up of Figure
6B. Shortly after hydroxyl ion
production has started, higher than normal positive ion concentration will
occur around the negative electrode from the positive ions drawn in by the
hydroxyl ion electric field, thereby shielding the hydroxyl ion electric field. The hydroxyl ions will not only keep
diffusing away from the negative electrode after their creation , they are also
actively being drawn toward the positive electrode. In Figure 6B the lines joining the two
electrodes are the electric field lines.
These lines represent the path that a negative or positive ion will
follow when traveling between the two electrodes, if they initially are located
on one of these lines and only the electric field is acting on the ion. The relatively shorter the line also
represent where the electric field is relatively the strongest and where the
ions therefore travel between the electrodes at the highest "drift"
velocity. As the hydroxyl ions are
generated at the negative electrode and bring in positive ions to shield their
electric field , they are also being guided along the field lines of Figure 6B
toward the positive electrode. However,
the hydroxyl ions bring (drag) much of their positive shielding ions with them. Furthermore, as the hydroxyl ions which were
generated on the shortest field lines travel away from the negative electrode
toward the positive electrode carrying much of their shielding positive ions
with them, other hydroxyl ions along with their positive ion shielding cloud
are transported onto the shortest field lines by ion density gradient
"pressures". The end or net
result is that sort of a expanding plum of hydroxyl ions along with their
positive shielding ions travel directly to the region of the positive electrode
by approximately the shortest path. The
cells surrounding the positive electrode thereby also experience a significant
increase in the positive ion concentration on the outside of their cell
membranes, along with a significant decrease in chlorine ion ( Cl- )
concentration from hydroxyl ion repulsion of chlorine ions. . These cells ( frog blood cells ) therefore go
through the same changes as the cells at the negative electrode, as was
described earlier. The positive hydrogen
ions generated at the positive electrode also react with the negative hydroxyl
ions to form water. However, the
hydrogen ion diffusion velocity is so much greater than that of the hydroxyl
ion that it rapidly diffuses out of the positive electrode region, thereby
allowing the plum of hydroxyl ions along with its shielding positive ions to be
drawn to the immediate vicinity of the positive electrode before the hydroxyl
ions and hydrogen ions significantly recombine.
So far I
have used simple hand waving qualitative electric field interaction arguments
to indicate how significant ion concentration changes can occur around
electrodes in saline solution to explain how cells can be effected by feeble
direct electric currents. To explain how
there is generated a positive ion concentration "wave" across the
chamber of Figure 6B is too complex and
beyond the scope and needs of this article.
Now that
we have arrived at a qualitative understanding of how the embryonic looking /
primitive dedifferentiated cells needed for blastema formation are created by
ionic conditions outside cell membranes, how do we apply this information in a
practical way ? All that is required is
for us to realize we need only scale up the results of the rat experiment of Figure 7A to that of
human size. For example, Figure 8 depicts a human finger which has been
amputated just above the first knuckle.
Platinum plated stainless steel acupuncture needles have been inserted directly
behind the amputation site a few days or so after the amputation. It is the ion concentrations that are the
missing critical factor. And the ion
concentrations required for blastema formation and maintenance should be about
the same for all mammals. So , all that
is required is to imagine that each acupuncture needle is taking care of one
rat arm section and then ask the question:
Approximately how many rat arm section crossectional areas do we have in
this amputated finger crossectional area ? That number is how many acupuncture needles
are required for the finger. Note that I
am using the same electrical circuit used in Figure 7A . That is the contact potential difference
between silver and platinum supplies the driving voltage and a high ohmage
carbon resistor limits / controls the current flow and therefore the hydroxyl
ion generation rate. Becker found that a
current in the neighborhood of two hundred nanoamps would work well in rat arm
regeneration experiments. Unfortunately,
there is a misprint or mistake on the optimal current required for rat forearm
regeneration stated on page 152 of Backer's book, THE BODY
ELECTRIC. It should state two
hundred nano - amps not one nano -
amp. Reference to the resistance range
used ( 106 to 108 ohms )
in the Figure on page 153 when placed into Equation
5 below shows the error. Also reference to the resistance ( 10 Meg
ohms ) used in cartilage regeneration experiments discussed on page 189 shows
the error. Another reason to expect the
current required in living tissue of
animals to be much larger than those required in Becker's frog blood cell experiment,
of Figure 6 A, is that that tissue has a blood supply to it. That blood supply system has associated with
it the continuous active pressure and diffusion driven transport of blood
plasma through the inter cellular spaces in the damaged tissue region. The hydroxyl ion generation rates are
therefore required to be much higher to compensate for the sweeping away /
dilution effects of the circulating blood supply in the damaged region.
Since the
contact potential difference between silver and platinum is about 2.3 volts, we
have from Ohm's Law :
Voltage
(V) =
Current (I) X Resistance (R) ;
Equation 5
V
= 2.3 volts and
the required current ( I )
is 200 x
10-9 amps = 2 x 10-7 amps
R
= V / I = 2.3 volts / 2 x 10-7amps =
11.5 x 106 ohms.
R is the required resistance to be in series with each acupuncture
needle. Note, this R value is right in
the middle of the resistance range ( 106 to 108
ohms ) used by Becker ,which was most
successful in his rat arm regeneration experiments. Becker would of had complete rat arm
regeneration, if he could of had the implant move along with the regrowth on
the arm as Smith did in his complete regrowth of a amputated frog limb.(5)
After the
blastema has formed and finger re growth has proceeded about one half a
centimeter all of the acupuncture needles should be reinserted into the finger
one half a centimeter in the forward direction.
In other words the acupuncture needles must be kept relatively close
behind the advancing blastema, until full regeneration has been achieved. What we have described here for the finger
can be done for the hand, foot, arm, leg, breast, etc. we must only make sure
to use enough needles evenly spaced apart with care taken to avoid the major
nerves, arteries, and veins. As the
crossectional area and therefore the thickness of the amputation site become
larger and larger, the length of the acupuncture needles must increase to stay
at least half the thickness at the amputation site and the current to each needle
must increase. The current increase
should be such that the current per unit length of the implanted needle remains
the same as that used at the original finger amputation site discussed above. By simply changing (lowering ) the value of
the high ohmage carbon resistors the current can be adjusted to the proper
value ( a constant current per unit length of implanted needle length ). As the regeneration process proceeds the
crossectional area of the advancing tissue will decrease. Less total current flow will be required to
maintain the proper ionic conditions.
Either or both a lessening of the number of needles used or a lowering
of the current is desirable. However,
when dealing with just a single finger one set of settings should do just fine.
The regeneration of body limbs should currently be a common
practice in the practice of medicine. It
is not, do to the rampant corruption at the highest levels of medical research
funding and therefore research control.
This fact was alluded to throughout Becker's book , and was discussed in
detail in his last "chapter" ;
Postscript: Political
Science. This corruption can not be over
stated. It is the driving reason behind
the high cost, high profit , ineffective ( for the patient / victim ) allopathic
medical system we are currently suffering under. For those of you who are becoming computer
literate , there is a Web Site on INTERNET managed by Walter W. Stewart <stewartw@helix.nih.gov> Try:
hffp://nyx10.cs.du.edu:8001/~wstewart/ This Site contains the complete text of
the Dingle Subcommittee Report on fraud and cover - up at the NIH.
In part 2
we will look at spinal cord regeneration, implications of ion concentrations
outside cell membranes for cancer treatment, and whole body regeneration /
rejuvenation in general.
IF YOU FOUND THIS ARTICLE OF REAL VALUE, PLEASE MAKE A HARD COPY WHILE STILL AVAILABLE.
References:
1) THE BODY ELECTRIC by Robert O. Becker, M.D.,
and Gary Selden; Published by William Morrow and Company, Inc.
2) Ibid
page 59
3) Ibid
page 113.
4) Ibid
page 143
5) Ibid
page 151
6) ELECTROGENIC
PROPERTIES OF THE Na
, K PUMP
by H. J. APELL,
J. MEMBRANE BIOL. 110.
103 - 114 (1989)
7) ACTIVATION
KINETICS OF SINGLE
HIGH - THRESHOLD CALCIUM CHANNELS
IN THE MEMBRANE
OF SENSORY NEURONS
FROM MOUSE EMBRYOS
BY P. G. KOSTYUK,
YA. M.
SHUBA, AND V. I. TESLENKO,
J. MEMBRANE BIOL.
110. 29 - 38 (1989)
8) SINGLE
CHLORIDE - SELECTIVE CHANNEL FROM
CARDIAC SARCOPLASMIC RETICULUM
STUDIED IN PLANAR
LIPID BILAYERS BY
ERIC ROUSSEAU, J. MEMBRANE BIOL. 110. 39 - 47 (1989)
9) Ca2+ - ACTIVATED K+
CHANNELS FROM CULTURED
RENAL MEDULLARY THICK
ASCENDING LIMB CELLS:
EFFECTS OF pH
BY MIGUEL CORNEJO, SANDRA E.
GUGGINO, AND WILLIAM
B. GUGGINO, J. MEMBRANE
BIOL. 110. 49 - 55 (1989)
10) MEMBRANE
POTENTIAL, ANION AND
CATION CONDUCTANCES IN
EHRLICH ASCITES TUMOR
CELLS BY IAN
HENRY LAMBERT, ELSE
KAY HOFFMANN, AND
FINN JORGENSEN, J.
MEMBRANE BIOL. 111.
113 - 132 (1989)
11) AN
ESSENTIAL 'SET' OF K+ CHANNELS
CONSERVED IN FLIES, MICE
AND HUMANS BY
LAWRENCE SALKOFF, KEITH
BAKER, ALICE BUTLER,
MANUEL COVARRUBIAS, MICHAEL
D. PAK AND
AGUAN WEI, TINS,
VOL. 15, NO. 5, 1992
12) PRIMARY
STRUCTURE AND FUNCTIONAL
EXPRESSION OF A
RAT G - PROTEIN - COUPLED MUSCARINIC
POTASSIUM CHANNEL BY
YOSHIHIRO KUBO, EITAN
REUVENY, PAUL A.
SLESINGER, YUH NUNG
JAN AND LILY
Y. JAN, NATURE ,
VOL.364,
13) THE
TIPS / TINS LECTURE: THE
MOLECULAR BIOLOGY OF
MAMMALIAN GLUTAMATE RECEPTOR
CHANNELS BY PETER
H. SEEBURG, TIPS - AUGUST 1993
(VOL.
14)
1 2 3
4 5 6 7 8 9 10 11
12
/ 05 / 95 PLEASE COPY / FAX
AND SHARE
SOME OTHER ARTICLES OF INTEREST
SYNERGISTIC ENERGY MEDICINE (NEW)
COLLECTED PAPERS ON RIFE TECHNOLOGY AND MORE
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}